
Written by: Anindita Sarkar
Edited by: Isabel Colon-Bernal, Christina Del Greco, & Alyse Krausz
My pet dog would be scratching and pawing me at roughly the same time every day without fail. My mother believes that he had a better sense of time than me and my sister. I would always convince my mom that the credit for his sense of time should be given to his internal biological clock. This internal biological clock, better known as the circadian rhythm, tunes different physiological processes in not just my dog but also other living beings, to a 24-hour cycle. That is why we tend to feel sleepy or hungry at almost the same time every day. Needless to say, this biological time-tracking system functions as a result of an intricate interplay of several genes and proteins. In fact, I work on one important member of this family of proteins, Rev-Erbβ, which is responsible for the proper upkeep of the circadian rhythm.
Rev-Erbβ is a transcriptional repressor and can, therefore, turn off or on protein synthesis by binding or not binding to a particular stretch of DNA aka gene sequence, respectively. This off and on mechanism allows Rev-Erbβ to regulate the expression of a core clock protein called BMAL1, which is also a transcriptional regulator.1,2 Together with a second protein BMAL1 regulates the cyclic expression of two important genes. This rhythmic process ultimately governs our sleep-wake cycle, hormonal oscillations, metabolism patterns, and many other physiological processes. The Rev-Erb family of proteins is also known to regulate the metabolism of lipids, inflammatory responses, generation of glucose, and formation of fat cells.1-4 However, Rev-Erbs need an indispensable ally (more scientifically known as a ligand)that facilitates its function as a transcriptional regulator. This ligand in question that binds to Rev-Erbβ is an iron-containing compound called heme, which gives blood its red color.5
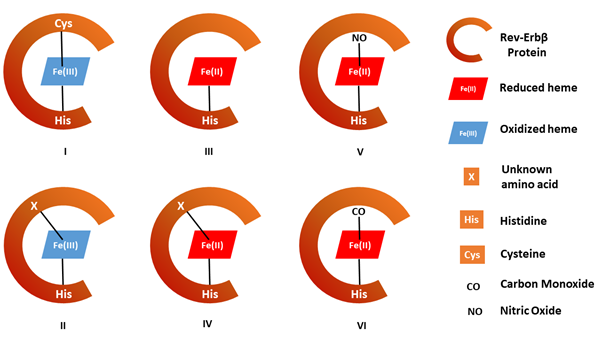
Heme is a versatile molecule. The iron center of the heme can exist in multiple oxidation states and/or can bind to different amino acids in Rev-Erbβ (Figure 1).6-8 Since heme-binding is crucial for Rev-Erbβ to function, we investigated the conditions under which the protein exists in the heme-bound state. We determined the redox potential of the heme bound to the protein. The redox potential precisely describes the propensity of a molecule (in this case heme) to lose an electron and undergo oxidation (Figure 1, Forms I and II) or gain an electron and undergo reduction (Figure 1, Forms III and IV). We found a very low redox potential for Rev-Erbβ, meaning that the heme should predominantly exist in the oxidized form in cells. Next, we performed experiments on bacterial cells engineered to express Rev-Erbβ protein verifying that the majority of the protein indeed exists in the oxidized form. The oxidized heme, in this case, is anchored to the protein by coordination through two amino acids: Histidine (His) and Cysteine (Cys) (Figure 1, Form I).9 Rev-Erbβ has a very high affinity for this particular form of heme, essentially making the protein a magnet that can pick up heme from the scarce free heme pools in cells.7 Furthermore, any cellular condition that alters how the heme is bound to Rev-Erbβ or the oxidation state of the bound heme leads to the dislodging of the heme from the protein, making it functionally impaired.
Another property of heme is that, in its reduced form, it interacts with several signaling gases like oxygen (O2), carbon monoxide (CO) and nitric oxide (NO).10 These gases interact with selective targets such as proteins or small molecules, like heme, to evoke certain physiological or biochemical changes in the cells, tissues, and/or the organism. The hemoglobin-mediated delivery of O2 to cells all over the body is a classic example of the interaction of gases at the heme center of a protein. Similar observations have been made for Rev-Erbβ, in which signaling gases can bind to the heme in the protein and evoke a cascade of downstream effects (Figure 1, Forms V and VI).9,11 This got us thinking: how does a protein that predominantly exists in the oxidized form suddenly get reduced to associate with these gases? To find this missing part of the jigsaw puzzle, we repeated our experiments of determining the redox potential of the protein, only this time in the presence of CO and NO. And guess what we found! These gases strongly favor the formation of the reduced heme. CO or NO can replace the Cys coordination of the heme (Figure 1, Form I) to yield the gas-bound reduced forms of the protein (Figure 1, Forms V and VI).


To visualize this process, imagine a tug of war between the oxidized and reduced forms of the protein (Figure 2). In the absence of the gases, the majority of the protein exists in the oxidized state. However, in the presence of CO or NO, the tables turn. Because the reduced form of the heme binds the gases so tightly, the protein is pulled from the oxidized to the reduced state. We observed this CO and NO-mediated Rev-Erbβ heme reduction in bacterial cells as well.
We believe this crucial observation can help explain an apparent anomaly for proteins that exist in the oxidized state but at the same time have been shown to function as gas-sensors like Rev-Erbβ. These findings also pave the way to explore the influence of signaling gases in mediating the circadian rhythm.
After explaining the importance of Rev-Erbβ and the complexity of the circadian rhythm to my mom, she was convinced that the molecular cascades were influencing my dog’s behavior. But she had a follow-up question: “How come I stay up so late at night – is my circadian rhythm out of whack ……?” I guess I’ll add that to the list of future research directions.
Check out Anindita’s published manuscript on this topic here!
Emoji from Pixabay.com
References:
1. https://doi.org/10.1038/nature11048
2. https://doi.org/10.1038/nrd4100
3. https://doi.org/10.1016/j.mce.2015.01.024
4. https://doi.org/10.1038/nature12209
5. https://doi.org/10.1074/jbc.R112.422642
6. https://doi.org/10.1074/jbc.M110.193466
7. https://doi.org/10.1074/jbc.M117.783118
8. https://doi.org/10.1074/jbc.M115.670281
9. https://doi.org/10.1371/journal.pbio.1000043
10. 10.1021/acs.chemrev.5b00018
11. https://doi.org/10.1101/gad.2064111

Anindita Sarkar is a postdoctoral fellow in Prof. S. W. Ragsdale’s laboratory at the Department of Biological Chemistry, University of Michigan. Her current research interest is studying the interactions of heme with core clock proteins that regulate the mammalian circadian rhythm. She completed her Ph.D. from the Tata Institute of Fundamental Research, India, under the guidance of Prof. Ankona Datta. Her graduate research work involved designing optical and MRI-based probes for essential metal ion, Manganese. Her hobbies include cooking, playing Sudoku, and reading about marine biology. She is passionate about communicating science to the non-scientific community. To know more about her work or to share some cool recipes you can connect with her on Linkedin.
