
Written and illustrated by: Jacquelyn Roberts
Edited by: Sarah Bassiouni, Sophia Hill, Austin Shannon, and Madeline Barron
Pathogenic bacteria like Salmonella, Shigella, Yersinia, and Escherichia coli can inject proteins into target cells with an extremely small hypodermic needle called a type III secretion system, or T3SS. The injected proteins are called “effector proteins”, as they elicit effects in the host (in this case human) cell. The cell membranes of both the bacteria and host cell prevent large molecules like effector proteins from simply drifting inside, so bacteria need specialized delivery systems like the T3SS to do it. These specialized delivery systems are very similar to a syringe and needle, and have been nicknamed injectisomes (Figure 1). However, injectisomes are 10 million times smaller than the average syringe. In addition to being incredibly small, these syringes are extremely specific in the cargo that they accept for transport. Bacteria generally use these machines to inject effector proteins that weaken the host cell, leading to easier bacterial infection. The mechanistic details of these molecular machines is of great interest to scientists for many reasons. Designing a specific inhibitor of the injectisome could render pathogenic bacteria harmless. On the other hand, engineering an injectisome for drug delivery could provide targeted biologic medicine to specific cells in the body.
Scientists’ first glimpses of the injectisome structure suggested that effector proteins travelled through the needle of the complex, an incredible feat considering that the “needle” of the injectisome is smaller in diameter than the average bacterial protein. This may seem impossible, but proteins are uniquely geared to perform this gymnastic act.

Proteins are first assembled like beads on a string, and then they are folded or knotted into their final form on the scale of angstroms (1Å = 0.0000000001m). These folds and knots can be loose or tight, but most importantly the overall shape allows the protein to be at a free energy minimum. This means that the proteins are more comfortable in their tangled-up form than they would be untangled. The genetic code, or DNA, of an organism dictates the order of the beads on the string. How the folding happens is less understood. The field of structural biology uses the shapes of folded proteins to answer biological questions. An average-sized protein produced by a bacterial cell like Salmonella can range from 30 – 60 Å in diameter. The inner diameter of the needle complex is only 20 Å. How would these effector proteins be transported through the needle?
Thomas Marlovitz and his lab at the Hamburg Center for Structural Biology investigated whether the effector proteins of Salmonella actually travelled through the needle-like projection of the bacterial cell. To do so, they looked at the injectisome in action. This may sound easy, but bacterial cells are incredibly small, and the injection of effector proteins occurs very quickly. So, the Marlovitz lab cleverly engineered effector proteins with large knots at the end to plug the needle of the injectisome mid “syringe-squeeze”. Then, they used a sophisticated technique known as single particle cryoEM to visualize the structure of the injectisome at near atomic resolution.
Basics of single particle CryoEM
CryoEM is a type of microscopy. Instead of a beam of light passing through the sample, it is a beam of electrons. The advantage of using an electron beam is that its wavelength is smaller than the wavelength of light. This allows the magnification of a sample more than 100,000x what we can see with the naked eye (or 1000x greater magnification than the highest setting on most light microscopes). The disadvantage of this electron beam is that it causes damage to the sample. This is where the cryo aspect of CryoEM enters the picture. To protect precious biological samples from the electron beam, they are flash frozen in ice. This keeps the sample hydrated, in a form like that found in living cells.
Single particle analysis is a way of analyzing CryoEM data to create 3D models of what was observed. It uses many different 2D images of the same particle (in this case, a protein) to reconstruct a model of that particle in 3D. In CryoEM the images collected are a projection of the particle, like a shadow. Imagine the particle was your hand, and the images collected were projections of light in the form of shadow puppets. A single shadow may not be enough to determine that the particle is in fact, a human hand. A talented shadow puppeteer may produce a very convincing “rabbit” shadow. If we collected shadows from all different directions of different “rabbits”, we could realistically average all the shadows to reconstruct a discernible human hand (Figure 2). The premise of single particle analysis is much the same. Injectisomes, being only hundreds of angstroms long, require magnification of more than 100,000x. The researchers in the Marlovitz lab collected their data at 112,998x magnification. They used over 82,000 different images of injectisomes that were extracted from 3 replicate data sets to reconstruct their 3-D model at a resolution of 2.4 – 4.5 angstroms. This is high enough resolution to discern the strands and knots of the protein string.

Transport in action
The Marlovitz lab began their study by manipulating one of the most well-studied Salmonella effector proteins. Inside the bacteria, the string of the effector protein is partially loose, and partially knotted in something like a slip knot. The untangled part of the protein is threaded into the injectisome to begin secretion. Once the knotted part of the protein is reached, it must be untangled to pass through the needle. This knot is short-lived; energy can quickly undo it. To get around this, the Marlovitz lab added a very strongly-tied knot to the end of the protein string. Importantly, this knot was opposite the end of the string that was fed into the needle of the injectisome. As a result, the string was halted mid injection, as the terminal knot could not be undone (Figure 3).
Using the plugged-up injectosomes and the power of single particle cryoEM, the researchers visualized a “gate” in the syringe that had two states: open and closed. When the effector protein enters the injectisome, parts within the injectisome “needle” rotate, leading to a larger opening inside of the injectisome. This allows the unfolded effector to travel through the central conduit of the injectisome.
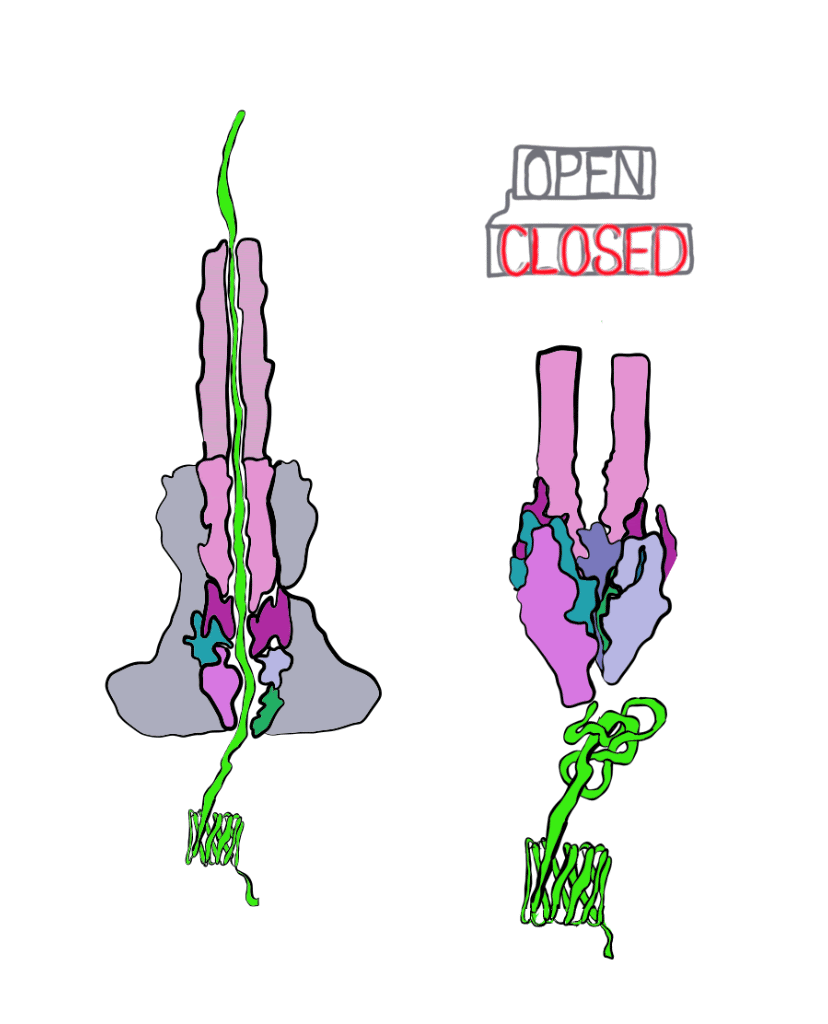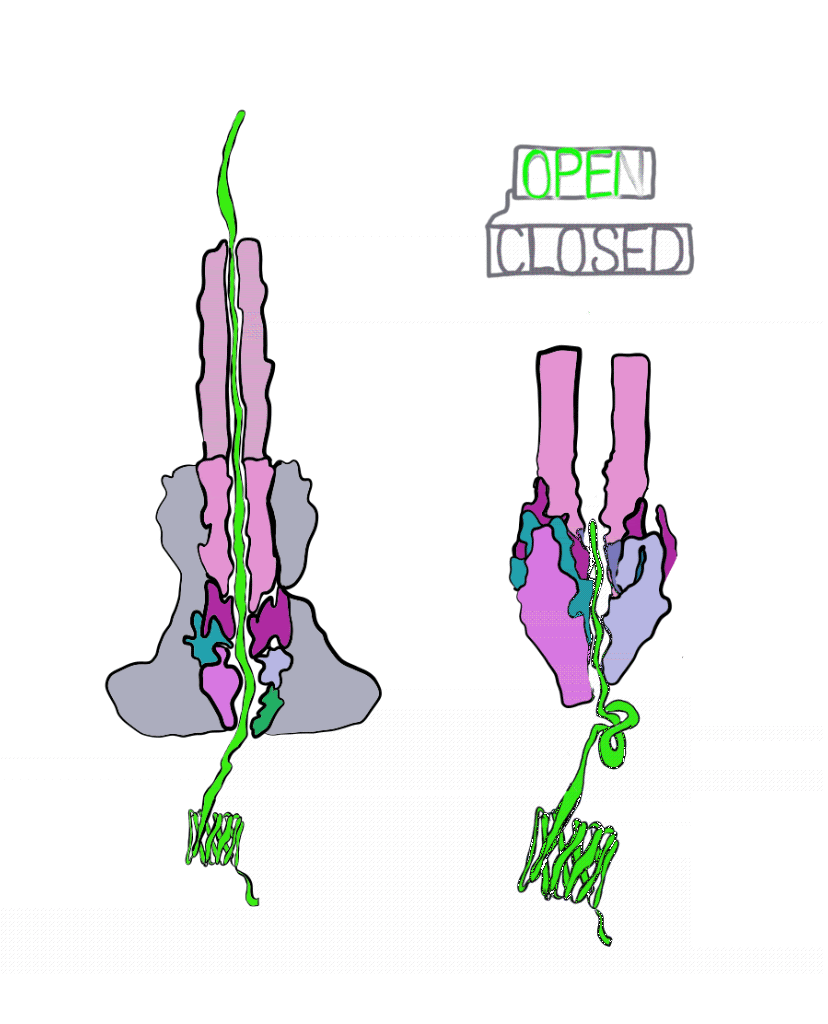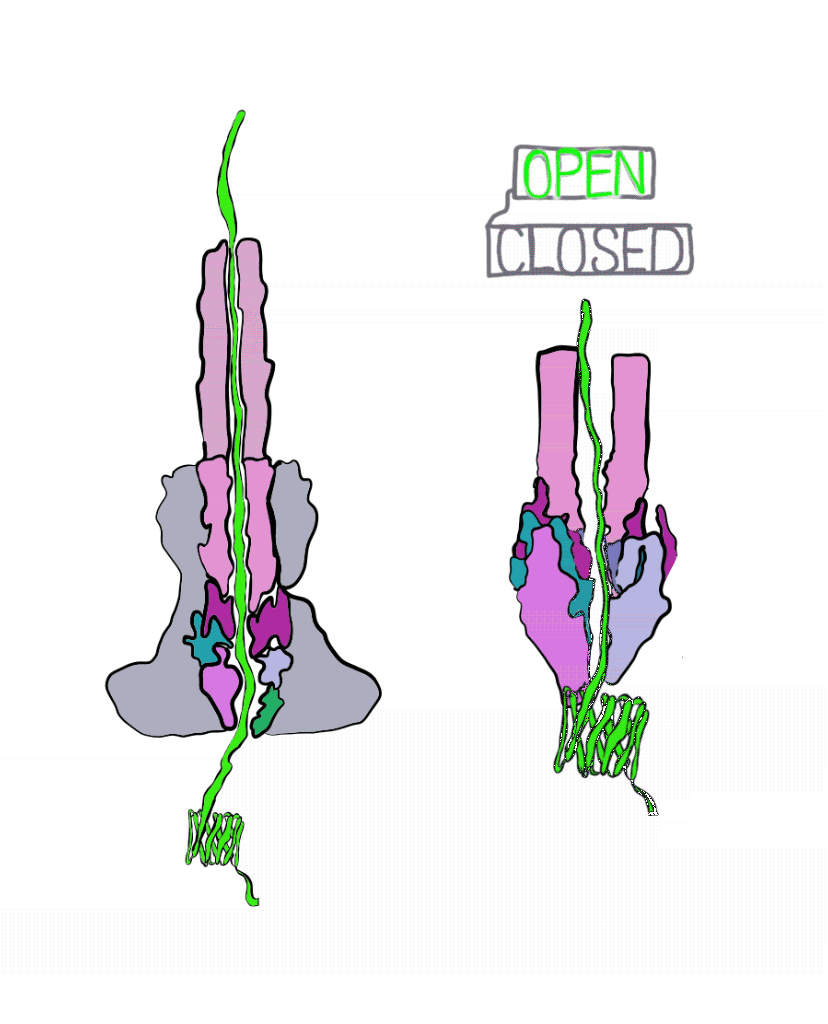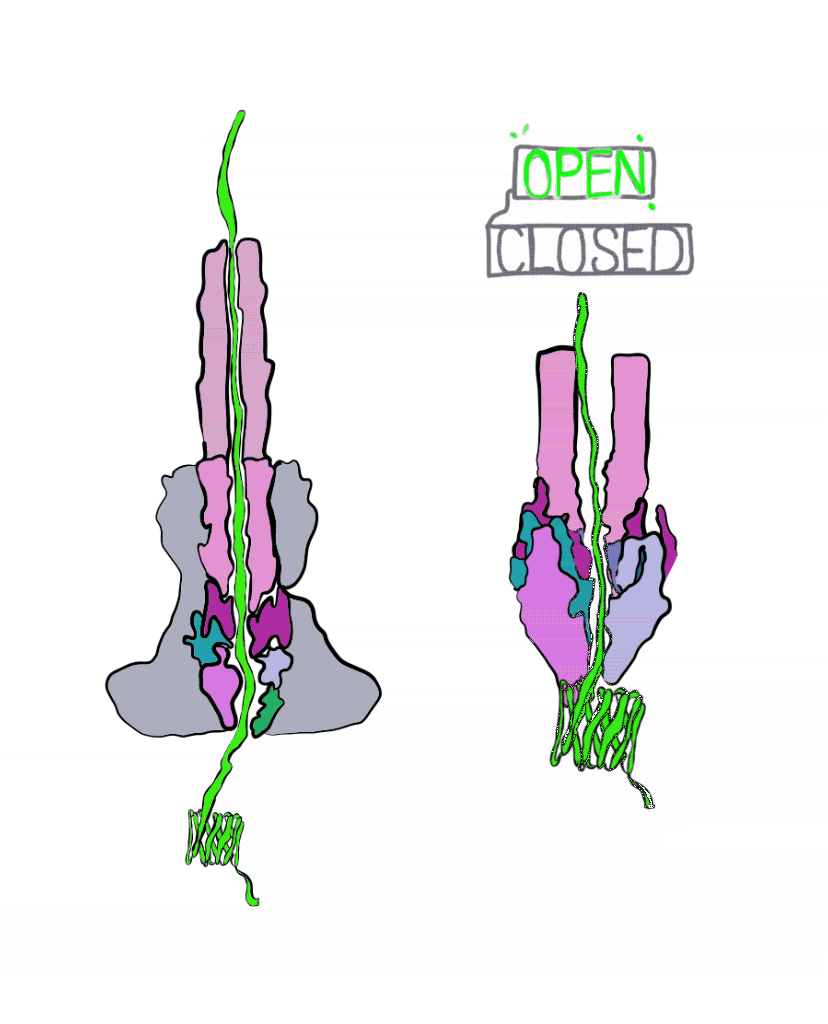
These details reveal the injectisome is more than a hypodermic needle; it is also a self-sealing hole in the membrane. Of course, questions remain unanswered. It is still unclear how exactly the injectisome recognizes the correct effector and rejects all others. Regardless, this study was integral to our understanding of bacterial secretion and paves the way for further study. Perhaps one day we will even be able harness the power of injectisomes for our own purposes, like the targeted delivery of drugs at the cellular level.

Jacquelyn Roberts is a Biological Chemistry Ph.D. Candidate in Melanie Ohi’s lab. Her research interests are focused on the structural biology of bacterial pore-forming toxins. Before starting her graduate work at the University of Michigan, she earned her Bachelors of Science at Eastern Michigan University in Ypsilanti. There she studied the effects of autophagy, or cellular self-eating, in Steven Backues’ lab. When not in lab, Jacquelyn likes to knit, draw, read science fiction, and cook savory dishes. She can be found on Twitter.

Wow! Thank you for such a great explanation of a very difficult to understand subject.
LikeLike