
Author: Molly A. Hirst
Edited by: Patricia Garay, Emily Glass, and Alyse Krausz
Most of us have a general understanding of what human sperm cells look like: a ball with a middle piece and a squiggly tail, right? But what about sperm from animals such as frogs, spiders, and mice? The short answer, as shown in Figure 1, is that they look totally different! In fact, sperm are actually the most morphologically diverse animal cell type known (seriously, check out Figure 1) (Lüpold & Pitnick 2018). This morphological diversity may seem counterintuitive at first; sperm have a singular function in fertilization and reproduction, so a mutation that alters the shape of sperm could potentially render them useless. This certainly can be the case, yet changes in sperm morphology between species, or even populations of the same species, are common.
Changes in sperm morphology can occur quite rapidly on an evolutionary timescale (Pitnick, Hosken, & Birkhead 2009, Pizzari & Parker 2009). The evolution of sperm traits and subsequent morphological differentiation can be explained by several well-supported evolutionary theories. One of the most prevalent and intuitive of these ideas is that sperm diversity (particularly in internally fertilizing animals) is the result of varying intensities of post-mating sexual selection pressures (Parker 1970, Lüpold & Pitnick 2018), where selection acts on components in the female and male reproductive tracts that are vital for proper reproduction (e.g., seminal fluid proteins that protect sperm on their journey through the female uterus).
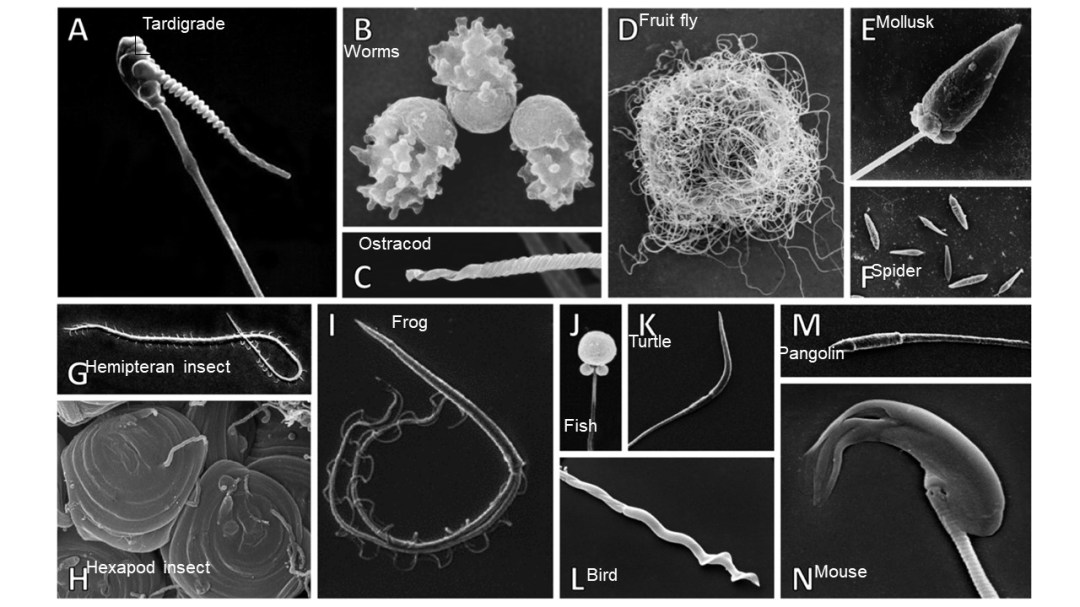 Figure 1: Sperm morphological variation represented by a single species of the following groups: A) tardigrades, B) worms, C) ostracods (a type of crustacean), D) fruit flies, E) mollusks, F) spiders, G) Hemipteran insects, H) Hexapod insects, I) frogs, J) fishes, K) turtles, L) birds, M) pangolins, and N) mice. (Image modified from Lüpold & Pitnick 2018).
Figure 1: Sperm morphological variation represented by a single species of the following groups: A) tardigrades, B) worms, C) ostracods (a type of crustacean), D) fruit flies, E) mollusks, F) spiders, G) Hemipteran insects, H) Hexapod insects, I) frogs, J) fishes, K) turtles, L) birds, M) pangolins, and N) mice. (Image modified from Lüpold & Pitnick 2018).
To understand how sperm cells evolve, we need to understand the basics of evolutionary theory. Evolution is the change in the genetics of a population over time. One of the ways in which this occurs is via selection, of which there are two “flavors”: natural selection and sexual selection. Natural selection favors certain compositions of genes that create traits that enhance the survival and reproduction of an individual (Darwin 1859, Arnold & Wade 1984, Nielson 2005). By surviving longer and producing more offspring, these individuals are more likely to pass their beneficial genes onto the next generation. Sexual selection occurs when evolution favors adaptations that only give individuals a reproductive advantage over others (Darwin 1871, Arnold & Wade 1984, Andersson 1994). This can occur either after mating, before mating, or both. A classic example of pre-mating sexual selection can be observed in peafowl. Males that have long tail feathers with many eyespots are more attractive to females than males with shorter tails and fewer eyespots (Figure 2) (Petrie & Halliday 1994). One hypothesis put forth to explain this pattern is the “sexy son” hypothesis, which states that a female will ideally choose a mate who will help her make the most and the strongest offspring (Huk & Winkel 2008). Another hypothesis is that these attractive male traits are “honest traits” that display a male’s health, which could be important for his future offspring’s viability, survivability, and reproduction (Loyau et al. 2005). Regardless of the hypothesis, there is a trade-off to having flamboyant, attractive tails: males with longer tails are more obvious to predators and may not be able to escape from danger as easily as their shorter-tailed counterparts, but they may be more likely to find a mate. This begs the question: if an attractive male with a long, showy tail is very successful at mating once, but won’t survive over the next several years, is the trade-off worth it? This example beautifully illustrates evolution via sexual selection by favoring traits that can increase a male’s reproductive success, but not his survivability.
 Figure 2: A male peacock displaying for a potential mate. (Photo by: Molly A. Hirst).
Figure 2: A male peacock displaying for a potential mate. (Photo by: Molly A. Hirst).
Although the example of pre-mating sexual selection in peafowl is quite obvious to the naked eye, sexual selection also acts on microscopic features of organisms that are important for processes that occur after mating, such as sperm morphology. One hypothesis of sexual selection theory that influences sperm morphological diversity is sperm competition (Parker 1970, Lüpold & Pitnick 2018). Sperm competition is, quite literally, competition between two or more sperm to reach and fertilize an egg. This competition can be 1) between two sperm from the same individual, 2) between sperm from two or more males of the same species, or even 3) between sperm from two or more males of different species!
- In the first scenario, this species is often designated as monogamous: one female mates with one male, and she does not mate with any other male. Thus, there is not much reason for sperm to evolve rapidly and drastically in this species because the male is guaranteed to be the father of the female’s offspring, regardless of how slow their sperm are.
- In the second scenario, the species has a high degree of female remating: the female mates with multiple males within a relatively short timeframe. This female remating provides the opportunity for intense sperm competition to occur because sperm from multiple males compete in utero to reach and fertilize an egg (Parker 1970). In this case, it is usually the male with the fastest, most numerous and long-surviving sperm that has the best chance to reach and fertilize an egg.
- The last scenario occurs when two species interbreed or hybridize. In this case, males not only compete with other males of the same species, but also with males from other species! This may be the subject of a future blog post… so stay tuned for more details! J
Knowing this, you can imagine that there is immense evolutionary pressure for males in the second scenario to have sperm traits that outcompete other male’s sperm to reach and fertilize an egg. You may also imagine that a male who produces sperm with more mitochondria, the part of the cell that produces energy required for sperm to swim, is likely to outcompete sperm with fewer mitochondria, and therefore less energy. Likewise, tail length can be indicative of faster-swimming sperm, so it’s likely that sperm with longer tails will outcompete those with shorter tails, et cetera. And if you’re wondering, the organism that claims the longest sperm relative to body size is a fruit fly (Drosophila bifurca), with sperm that are a whopping ~5.8 centimeters long when unwound (Lüpold et al. 2016)! Because these sperm are so large (for such a tiny organism!), each sperm is wound up into a ball-like shape and very few sperm (relative to humans, for example) are made at a given time (Lüpold et al. 2016).
We know one simple yet complex reason why sperm morphology differs between organisms- it’s all about sexual selection! However, there is still a lot we do not know about the genetic or environmental basis of sperm morphological diversity. In general, evolutionary biologists are trying to answer the question: why and how are sperm so different between species?
As a sperm evolutionary biologist and sperm enthusiast, I find that I enjoy a slightly modified version of one of Charles Darwin’s famous quotes: “from so simple a beginning, endless [sperm] forms most beautiful and most wonderful have been, and are being, evolved.” Just as all animals on Earth are diverse (from flies to frogs to humans), so, too, are their sperm.
 Molly A. Hirst is a graduate student at the University of Michigan in the department of Ecology and Evolutionary Biology (EEB). She is co-advised by Liliana Cortés-Ortiz and Trisha Wittkopp. Molly grew up in Michigan and forgot to ever leave. She has four degrees that she earned at various institutions in Michigan (A.S., B.S., B.A., M.S.) and is currently finishing her PhD in EEB at the University of Michigan. Her research interests include sperm morphology in primates and how hybridization (interbreeding and genetic admixture) between primate species affects sperm in hybrid individuals. She is interested in all things natural history including learning about museum collections and traveling the world to find cool animals and learn about various cultures. Her hobbies include travel and nature photography, sudoku, raising succulents and her two cats, reading, spoiling her nephew, constantly learning about self-development, and enjoying a good cup of coffee.
Molly A. Hirst is a graduate student at the University of Michigan in the department of Ecology and Evolutionary Biology (EEB). She is co-advised by Liliana Cortés-Ortiz and Trisha Wittkopp. Molly grew up in Michigan and forgot to ever leave. She has four degrees that she earned at various institutions in Michigan (A.S., B.S., B.A., M.S.) and is currently finishing her PhD in EEB at the University of Michigan. Her research interests include sperm morphology in primates and how hybridization (interbreeding and genetic admixture) between primate species affects sperm in hybrid individuals. She is interested in all things natural history including learning about museum collections and traveling the world to find cool animals and learn about various cultures. Her hobbies include travel and nature photography, sudoku, raising succulents and her two cats, reading, spoiling her nephew, constantly learning about self-development, and enjoying a good cup of coffee.

Thanks for sharing this well written article which is specially designed for online readers.
LikeLike